Taoyuan Ran
Abstract. This research will analyse the different roles and interrelationships of PC1 and PC2 enzymes in obesity and diabetes. There are significant differences in substrate specificity and function of the two enzymes during metabolism. PC1 is mainly involved in the negative feedback regulation of insulin signals, while PC2 can play a key role in adipocyte differentiation and energy metabolism. Based on this, the regulation of PC1 and PC2 enzymes will provide an effective scheme for the regulation of related metabolic diseases. In addition, this research will explain how dopamine and hormones secreted by the pituitary gland (GH, ACTH) act on the mRNA expression of PC1 and PC2 enzymes, thereby affecting insulin secretion. Inhibition of PC1 activity can improve insulin resistance, thereby alleviating the symptoms of type 2 diabetes. Conversely, enhancing PC2 activity may help reduce body fat accumulation and combat obesity. Understanding the differences between these two enzymes is important for developing new therapeutic strategies.
1 Introduction
The research on the mechanism of insulin synthesis and secretion has been paid more and more attention due to the increasing incidence of diabetes. Insulin can be used to regulate blood sugar levels and is a polypeptide hormone secreted by beta cells containing 51 amino acid residues, which consists of three separate functional regions: two identical chains (A and B chains), connected in the middle by a longer and irregular C chain. In its natural state, proinsulin is first captured by signal recognition particles (SRP) on the rough endoplasmic reticulum and subsequently further processed in the Golgi complex. At this stage, proinsulin is cut by two specific protein convertases, PC1 and PC2, to form the correct amino acid sequence and transported to the cell surface through vesicles for release. PC1 first cuts off a specific site of proinsulin to produce an intermediate, a C-peptide, along with a small molecule called a PC2 activator. This small molecule then directs the second cleavage, the result of PC2 acting on the other end of the C-peptide, to complete the entire insulin maturation process. At this point, the proinsulin is converted and processed into active insulin.
In recent years, the mechanism of PC1 and PC2 enzymes in insulin processing is investigated extensively [1]. For example, gene knockout technology can be used to observe changes in mouse models lacking PC1 or PC2. The results suggest that PC1 is necessary for hormone processing, and on the other hand, although PC2 can play a key role in some hormone processing, its absence does not always cause a distinct phenotype. In addition to their role in normal physiological conditions, PC1 and PC2 also play a key role in responding to pathological conditions such as diabetes. An epidemiological survey based on human subjects has shown that certain types of genetic variants can affect PC1/PC2 expression or alter their cutting efficiency, making individuals more susceptible to type 2 diabetes.
The proinsulin processing theory is the biochemical explanation of how insulin is converted from its precursor form, proinsulin, into its active form. The process begins with the transcription and translation of the insulin gene in the rough endoplasmic reticulum (RER) of pancreatic beta cells to insulin is activated and becomes capable of binding to insulin receptors on target tissues to regulate glucose metabolism. Dysregulation of proinsulin processing can result in the development of diabetes mellitus or other endocrine disorders. Understanding this process is therefore crucial for the diagnosis and treatment of these conditions.
In the background of proinsulin processing theory, this research will focus on the functional differences between PC1 and PC2 and resultant effects on insulin processing. Although both belong to the family of membrane-bound serine proteinases, the tissue distribution and function of each expression are different. Among them, PC1 is mainly responsible for activating the double-skin circular proinsulin produced by E. coli into biologically active linear insulin. PC2 is considered to be a key factor in maintaining the homeostasis of insulin metabolism at normal plasma level and to further investigate their association with related diseases. An abnormally increased immature form of proinsulin due to mutations in the PC1 gene has been found in patients with inherited hyperinsulinemia. At the same time, there are also studies that suggest that PC2 defects may cause the development of similar disorders or exacerbate symptoms.
2 The differences between PC1 and PC2 enzyme function
PC1 and PC2 are verified to be significant roles for cleavage. However, their specific function that is, the cutting site and rate are not the same. Although PC1 plays a larger critical role than PC2 in the conversion of proinsulin, this does not mean that PC2 is less functional, in fact PC2 can play a key role for the processing of other pre-islet hormones, particularly the pre-conversion of glucagon to glucose [1]. PC1 removes excess sequence at the C-end, while PC2 precisely cuts an amino acid junction between the B and A chains in the middle. More specifically, PC1, a protease located in the Golgi apparatus, recognizes and cuts a polypeptide sequence containing R-X-K/R-R, where X can be any amino acid. PC1 first cuts the C-peptide region of proinsulin, separating the B chain from the A chain to form an intermediate consisting of the B chain and the C-peptide. This cleavage site is usually an RXXR sequence. PC2 then performs a cleavage at the N-terminal of the C-peptide, removing the C-peptide and leaving the mature insulin molecule. This cleavage site is an RX(R/K)R sequence, where R or K in parentheses is the cleavage site of PC2.
This difference leads to the difference in cleavage efficiency between the two. In general, PC1 usually exhibits a higher catalytic rate constant because of its simple and well-positioned task, while PC2 has a slower catalytic rate because it must accurately identify a less obvious site [2]. The results show that the expression level of PC1 was much higher in β cells than in non-β cells, while the expression level of PC2 was lower. Despite these reservations, it was perfectly valid to compare PC1 or PC2 levels and their proportions. The results show that PC1 levels in beta cells were more than 20 times higher than in non-beta cells. In contrast, PC2 levels in non-beta cells were about twice as high as those in beta cells. This suggests that PC1 is clearly more effective in insulin treatment. In addition, in the transformed beta cells, PC1 levels were abnormally low and mouse proinsulin conversion was significantly impaired, while in the AtT20 cells, it was fast and effective, with very high PCI levels and extremely low PC2 amounts. For human insulin initially said, the situation may be different.
In accord with insulin processing theory, PC1 and PC2 enzymes play as key factors of insulin activating. Although both can promote the activation of secreted proteins, they show different preferences and efficiencies in practice. For example, PC1 prefers a shorter hydrophobic side chain as an amino acid residue at the P1 position. In contrast, PC2 prefers residues with long hydrophobic side chains or polar heads as the best choice for this location. In the case of natural substrates, changing the type and sequence of amino acids near the cutting site on the substrate had a significant effect on the catalytic effect of both. Especially, as the transition from pre-proinsulin to proinsulin to mature insulin, this difference becomes particularly pronounced when it comes to key steps that directly affect product yield.
Under this substrate specific condition, PC1 and PC2 are controlled by multiple regulatory mechanisms at the molecular level, leading to differences in spatial and temporal expression patterns. Increasing the expression level of PC1 while keeping the level of PC2 unchanged will lead to the accumulation of more immature forms, because excessive PC1 leads to inappropriate selection of cutting sites. On the contrary, increasing the expression of PC2 will speed up the processing speed but may sacrifice some accuracy. This suggests a dynamic balance and helps to understand why product purity issues often arise when insulin is manufactured on a large scale [3].
3 Regulation of PC1 and PC2 enzymes
As a widespread central nervous system transmitter, dopamine also has regulatory functions in peripheral tissues. Dopamine can influence the function of islet beta cells through its receptor subtype, further regulating the activity of PC1/PC2 enzymes. Activation of D1-like receptors promotes the activity of adenylate cyclase, increases the level of cAMP, and then activates the PKA pathway. This change may enhance the activity of the PC1/PC2 enzyme, promoting insulin processing and release. Moreover, activation of D2-like receptors reduced cAMP levels, and led to inhibition of PKA pathway. This may affect the phosphorylation status of PC1/PC2 enzymes, thereby altering their activity and ultimately affecting insulin processing and releasing [4].
Specifically, activation of D1-like receptors can promote adenylate cyclase activity and increase cAMP levels, which in turn activates the PKA pathway. This change may enhance the activity of the PC1/PC2 enzyme, promoting insulin processing and release. D2-type dopamine receptor agonists can inhibit the catalytic activity of PC1/PC2 and reduce insulin release. This suggests a potential strategy to use drugs to interfere with dopamine signaling pathways to fine-tune blood sugar control. A good example is that the expression of PC1 and PC2 genes in the anterior lobe is significantly affected by haloperidol and bromocriptine. Haloperidol causes a great influence on PC2 mRNA levels about an increase of 1 to 8 times, but it only impacts the mRNA expression level of PC1 negligibly.
The pituitary gland releases a number of signaling substances, such as neurotransmitters and polypeptide hormones, which can reach the islet beta cells through the blood circulation and directly or indirectly regulate the activity of PC1 and PC2 [5]. For example, GH can stimulate the proliferation and function maintenance of islet beta cells. GH can increase the expression of PC2 gene by activating JAK/STAT pathway, thus improving the efficiency of insulin processing. Also, GH stimulates the liver to produce insulin-like growth factor (IGF), which has been shown to directly or indirectly enhance PC2 expression and activity. Specifically, IGF signaling pathway can promote PC2 gene expression by up-regulating MafA transcription factor activity through PI3K-Akt pathway. Dexamethasone can down-regulate POMC mRNA levels in AtT-20 cells. AtT-20 cells have high expression of PC1 mRNA but less expression of PC2 mRNA, very similar to anterior corticotropic cells. Dexamethasone treatment rapidly reduced PC1 and PC2 mRNA levels in AtT-20 cells by about 50%.
Additionally, ACTH can significantly up-regulate the transcription and translation levels of PC1/PC2 genes. Specifically, corticotropin is a peptide hormone and its main role is to stimulate the secretion of the adrenal cortex. ACTH can not only promote the secretion of glucocorticoid, but also up-regulate the expression level of PC1, enhance its enzyme activity, and accelerate the conversion of proinsulin to insulin [6].
4 Related diseases
PC1 plays a key role in the oxidation of fatty acids. It promotes fat accumulation by cutting and activating specific substrates. When PC1 activity is reduced, it leads to fatty acid accumulation. Loss of PC1 significantly reduces the rate of β-oxidation of fatty acids. When the PC1 gene was knocked out in mice, the animals were more prone to weight gain and insulin resistance. The role of PC2 in metabolic diseases is more complex and diverse than that of PC1 [7, 8], as shown in Fig. 1. It not only participates in lipid metabolism, but also can play a key role in inflammatory response and cell proliferation. PC2 can promote adipocyte differentiation and lipid storage. When PC2 is overexpressed, it may exacerbate the development of obesity. However, moderate up-regulation of PC2 activity can effectively increase energy expenditure and prevent the occurrence of obesity.
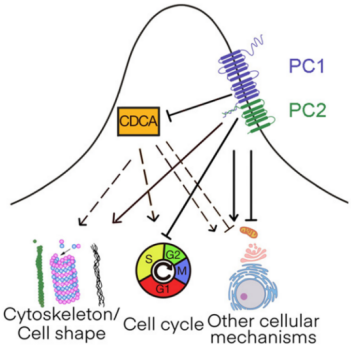
Fig. 1. PC-based signaling control by multiple responding nodes [8].
In addition, PC1 can be used for the regulation of glucose metabolism [9]. Decreased PC1 activity leads to impaired glucose tolerance and elevated blood sugar levels. The expression level of PC1 is significantly reduced, resulting in insufficient insulin secretion or increased insulin resistance. The dysfunction of PC1 directly affects the synthesis and release of insulin, thus aggravating the symptoms of diabetes. A study in a mouse model showed that knocking out the PC1 gene leads to a severe diabetic phenotype, demonstrating its importance in maintaining normal glucose homeostasis. Therefore, enhancing the activity of PC1 is expected to improve the symptoms of diabetic patients. In contrast, the role of PC2 is more complex. For example, it can be used to regulate the production of active forms of other peptides such as GLP-1. PC2 defects can lead to the inability of GLP-1 to activate effectively, which in turn reduces insulin secretion and leads to hyperglycemia. PC2 is not only a key factor in insulin processing, but also plays a crucial role in intestinal L cells [10].
5 Conclusion
In summary, although PC1 and PC2 are both important members of the protein tyrosine phosphatase family, they show significant differences in function, expression regulation mechanism and pathophysiological effects. Due to substrate specificity, PC1 and PC2 differ in their preference and efficiency in processing and activating proinsulin, while PC1 generally exhibits higher catalytic efficiency. The conclusion of the discussion on the regulatory mechanism shows that dopamine and hormones secreted by the pituitary gland can effectively promote mRNA expression and further increase insulin secretion. When the scale of insulin secretion expands, obesity and diabetes will be effectively inhibited and treated.
References
1. A. Ramzy, T. J. Kieffer, Altered islet prohormone processing: a cause or consequence of diabetes?.Physiological reviews, 102(1), 155-208 (2022)
2. M. Neerman-Arbez, V. Cirulli, P. A. Halban, Levels of the conversion endoproteases PC1 (PC3) and PC2 distinguish between insulin-producing pancreatic islet β cells and non-β cells. Biochemical Journal, 300(1), 57-61 (1994)
3. P. A. Halban, Proinsulin processing in the regulated and the constitutive secretory pathway. Diabetologia, 37, S65-S72 (1994)
4. R. Day, M. K. Schafer, S. J. Watson, et al. Distribution and regulation of the prohormone convertases PC1 and PC2 in the rat pituitary. Molecular Endocrinology, 6(3), 485-497 (1992)
5. X. Zhu, A. Zhou, A. Dey, et al. Disruption of PC1/3 expression in mice causes dwarfism and multiple neuroendocrine peptide processing defects. PNAS, 99(16), 10293-10298 (2002)
6. L. Jin, E. Kulig, X. Qian, et al. Distribution and regulation of proconvertases PC1 and PC2 in human pituitary adenomas. Pituitary, 1, 187-195 (1999)
7. S. Ozawa, H. Katsuta, K. Suzuki, et al. Estimated proinsulin processing activity of prohormone convertase (PC) 1/3 rather than PC2 is decreased in pancreatic β-cells of type 2 diabetic patients. Endocrine Journal, 61(6), 607-614 (2014)
8. L. Luo, S. Roy, L. Li, M. Ma, Polycystic kidney disease: Novel insights into polycystin function. Trends in Molecular Medicine, 29(4), 268-281 (2023)
9. S. R. Islam, S. K. Manna, Identification of glucose-independent and reversible metabolic pathways associated with anti-proliferative effect of metformin in liver cancer cells. Metabolomics, 20(2), 29 (2024)
10. K. Baafi, J. C. March, Harnessing gut cells for functional insulin production: Strategies and challenges. Biotechnology Notes, 4, 7-13 (2023)
Leave a Reply
You must be logged in to post a comment.